Rebecca E. Dey and Skyler T. Carpenter, authors
Abstract
Cabbage white butterflies, Pieris rapae, are agricultural pests that lay their eggs on plants of the Brassicaceae (cabbage) family. Upon hatching, the caterpillars then feed on the plants, causing contamination and yield reduction. Female butterflies may identify suitable host plants through the detection of various chemicals produced by the plants. Of particular interest are phenolic acids because they are found in high concentrations in Brassicaceae plants, yet little is known about how they may influence egg-laying (ovipositional) behavior. This study evaluated the potential of four phenolic acids to serve as ovipositional stimulants or deterrents for Pieris rapae. Treatments of phenolic acids and deionized water controls were applied to cabbage disks that were equal in leaf surface area and color. Each treated disk was matched with a deionized water control and placed within an experimental enclosure with a single mated female. After 24 hours, the number of eggs laid on each disk was observed and recorded. Two of the phenolic acids (caffeic and sinapic) deterred oviposition, one (cinnamic acid) was a stimulant, and one (vanillic acid) had no significant effect. The combined results of this study and previous research suggest that it may not be possible to make broad generalizations about the effects of phenolic acids on ovipositional choices and underscore the importance of assessing individual chemicals. Understanding what chemicals, and other factors, serve as stimulants or deterrents for oviposition may provide valuable information for avoiding or minimizing the negative effects of Pieris rapae on agricultural crops.
Introduction
Cabbage white butterflies (Pieris rapae) lay their eggs (oviposit) on plants in the Brassicaceae family, such as cabbage, turnips, and kale (Richards 1940). When the eggs hatch, the caterpillar larvae consume the host plant and use it as their primary food source. The nutritional value of the host plant is therefore critical for the success and development of the caterpillars. In order to increase the female butterfly’s fitness, it is advantageous for her eggs to be laid on the best host plants possible. It is thought that females identify suitable plants using visual and olfactory cues (Renwick and Radke 1988; Schoonhoven et al., 2005), including detection of various chemicals (plant secondary metabolites) that are produced by the host plants.
Previous studies have shown that P. rapae oviposition is affected by the presence of glucosinolates and cardenolides in plants of the Brassicaceae family. Glucosinolates were found to stimulate oviposition (Renwick and Radke, 1988), whereas cardeolides were found to deter oviposition (Renwick et al. 1989). However, other classes of chemicals are present in Brassicaceae plants, many of which could also have an effect on the oviposition of
P. rapae. The secondary metabolites of particular interest are phenolic acids, because they are found in high concentrations in some Brassicaceae plants (Mattlia and Hellstrom, 2007), yet little is known about the role they play in ovipositional choices. The main objective of this study was to evaluate the potential for phenolic acids to serve as ovipositional stimulants or deterrents for Pieris rapae.
Methonds
Wisconsin Fast Plants®, Brassica rapa, were planted in Styrofoam planting units using vermiculite as a soil-like medium for plant growth. The planting unit consisted of four compartments. One plant seed and two fertilizer pellets were placed in each compartment. The planting units were placed on a watering and nutrient tray reservoir. Through a felt-wick system, the plants were provided with a continuous nutrient and water supply. Plants were then placed in a growth chamber where the environment was controlled. The chamber was maintained at 26° C and provided continuous light. The chamber had a combination of fluorescent and incandescent lighting (1520 Footcandles). After 1–2 weeks of plant growth, we placed P. rapae eggs onto the leaves of the plants. The eggs were purchased from Carolina Biological Supply Company. After the caterpillars had grown to a proper size, we removed them from the Brassica plants and placed them into plastic cups with lids that provided adequate ventilation for their survival. Slices of store-bought cabbage were placed in the cups for the caterpillar’s primary food source. The cups were cleaned daily and a new food supply was given until their early stages of pupation. Caterpillars were placed onto a sheet of paper inside their enclosures to continue pupation (roughly 1–2 weeks before emergence occurred). Males were identified by one spot on their wings, whereas females had two spots on their wings. After butterfly emergence and the allowance of adequate time for mating (1–3 days) females were selected for the experiments.
Treatments of 1 mM phenolic acids and deionized water (DI H20) controls were applied to cabbage disks that were equal in color and leaf surface area (6.4 cm diameter disks). The phenolic acids that were tested individually in this study were sinapic (n=15), vanillic (n=10), caffeic (n=22), and cinnamic (n=13) acids. Each phenolic acid–treated disk was matched with a DI H20 control disk and placed at opposite ends of an experimental mesh enclosure (38 cm wide x 61 cm tall cylinder). The female butterflies were able to choose equally between the cabbage disks. One female butterfly was placed in each enclosure.
Females were given 24 hours to lay eggs. The choice of female oviposition and the number of eggs laid on each disk were observed and recorded. Data were analyzed with the JMP® statistical program. Paired t-tests were performed for each phenolic acid experiment to evaluate differences between the number of eggs that a female laid on phenolic acid treatment and control cabbage disks.
Results and Discussion
The complex chemicals produced by plants, called plant secondary metabolites, are thought to play a major role in the orientation of insects to their host plants and in the avoidance of laying eggs on plants that would be unsuitable for the development of their larvae. However, much is still unknown about the specific chemicals that are responsible for this attraction or deterrence. This study examined the effects of phenolic acids on the ovipositional choices of Pieris rapae (cabbage white butterflies). Phenolic acids are aromatic compounds that are ubiquitously produced, but are found in particularly high concentrations in the Brassicaceae host plants of P. rapae (Mattlia and Hellstrom, 2007).
In this study, three of the four phenolic acids showed a significant effect (p < 0.05) on Pieris rapae oviposition. Caffeic and sinapic acids both acted as deterrents; the butterflies laid more eggs on the cabbage disks containing deionized water than they did on the disks treated with either acid (Figure 1). 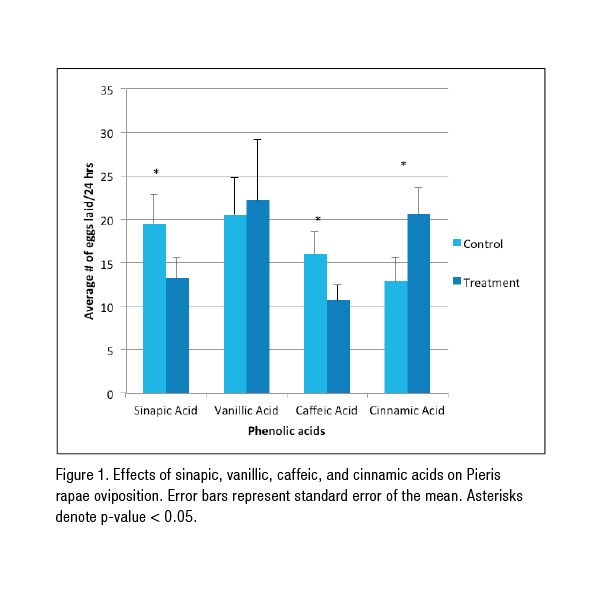 Cinnamic acid acted as a stimulant; the butterflies preferred the cinnamic acid–treated cabbage disks to the deionized water control disks (Figure 1). Vanillic acid acted as neither a stimulant nor a deterrent. It had no significant effect on oviposition (Figure 1).
Cinnamic acid acted as a stimulant; the butterflies preferred the cinnamic acid–treated cabbage disks to the deionized water control disks (Figure 1). Vanillic acid acted as neither a stimulant nor a deterrent. It had no significant effect on oviposition (Figure 1).
Previous research done by Walker et al. (2014) found that p-coumaric and ferulic acids had a significant stimulatory effect on
P. rapae oviposition on intact Brassica rapa plants. Walker et al. (2014) also found that protocatechuic and salicylic acids did not significantly affect oviposition.
Taken together, the results of this study and that of Walker et al. (2014) provide evidence that phenolic acids have a broad range of effects on P. rapae oviposition. The effects range from stimulatory, to deterrent, to having no effect. Results from both studies show that five of the eight phenolic acids tested had a significant effect on oviposition. Walker et al. (2014) suggested that the variability in effects of phenolic acids may be influenced by the chemical structure of the compounds. However, in examining how the chemical structure of phenolic acids relate to effect (deterrent, stimulatory, or no effect) with the additional acids included in our study, no consistent trends were observed. Cinnamic, caffeic, p-coumaric, and ferulic acid all have similar structures; however, three caused a stimulatory effect and one caused a deterrent effect. Differential responses of females to the types of phenolic acids suggest that it may not be possible to make broad generalizations about their effects on ovipositional decisions and underscores the importance of assessing individual chemicals.
There are many factors besides plant secondary metabolites that might have an effect on P. rapae oviposition. The studies of Traynier (1986) have shown that P. rapae associate visual cues with the presence of possible stimulants and suggest that the size and coloration of Brassicaceae plants influence
P. rapae oviposition. The nutritional value of Brassicaceae plants could also play a role in P. rapae ovipositon (Walker et al., 2014). P. rapae female butterflies want to lay their eggs on the healthiest plants possible to secure the survival of their young and increase their overall fitness. Additionally, young Brassicaceae plants contain lower levels of phenolic compounds and may differ in the concentrations of other plant secondary metabolites. This may contribute to P. rapae’s preference for younger Brassicaceae plants, since sensory perception of phenolic acids has been demonstrated in Pieris caterpillars (van Loon 1990).
Pieris rapae caterpillars can cause considerable damage to Brassicaceae agriculture plants by large consumption rates and by contamination with excrement, which in turn makes the plants unmarketable (Hern et al., 1996). Understanding what chemicals, and other factors, serve as stimulants or deterrents for female oviposition may provide valuable information for avoiding or minimizing these negative effects of their larvae.
Literature Cited
Hern, A., G. Edward-Jones, and R.G. McKinlay. 1996. A review of the pre-oviposition behaviour of the small cabbage white butterfly, Pieris rapae (Lepidoptera: Pieridae). Annals of Applied Biology 128:349-371.
Mattila, P. and J. Hellström. 2007. Phenolic acids in potatoes, vegetables, and some of their products. Journal of Food Composition and Analysis 20:152–160.
Renwick, J. A. A. 1989 Chemical ecology of oviposition in phytophagous insects. Experientia 45: 223–228.
Renwick, J. A. A. and C. D. Radke. 1988. Sensory cues in host selection for oviposition by the cabbage butterfly, Pieris rapae. Journal of Insect Physiology 34:251–257.
Renwick, J. A. A., C. D. Radke, and K. Sachdev-Gupta. 1989. Chemical constituents of Erysimum cheiranthoides deterring oviposition by the cabbage butterfly, Pieris rapae. Journal of Chemical Ecology 15:2161-2169.
Richards, O.W. 1940. The biology of the small white butterfly (Pieris rapae), with special reference to the factors controlling its abundance. Journal of Animal Ecology 9: 243-288.
Schoonhoven, L.M., J.J.A. van Loon, and M. Dicke. 2005. Insect-Plant Biology. Oxford: Oxford University Press.
Traynier, R.M.M. 1986. Visual learning in assays of sinigrin solution as an oviposition releaser for the cabbage butterfly, Pieris rapae. Entomologia Experimentalis et Applicata 40:25–33.
van Loon, J.J.A. 1990. Chemoreception of phenolic acids and flavonoids in larvae of two species of Pieris. Journal of Comparative Physiology 166:889-899.
Walker, K.S., J.L. Bray, M.E. Lehman, and A.J. Lentz-Ronning. 2014. Effects of host plant phenolic acids and nutrient status on oviposition and feeding of the cabbage white butterfly, Pieris rapae. BIOS 85:95-101.